Archives
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-11
- 2018-10
- 2018-07
-
After ischemia expression of transcription factors including
2022-03-03
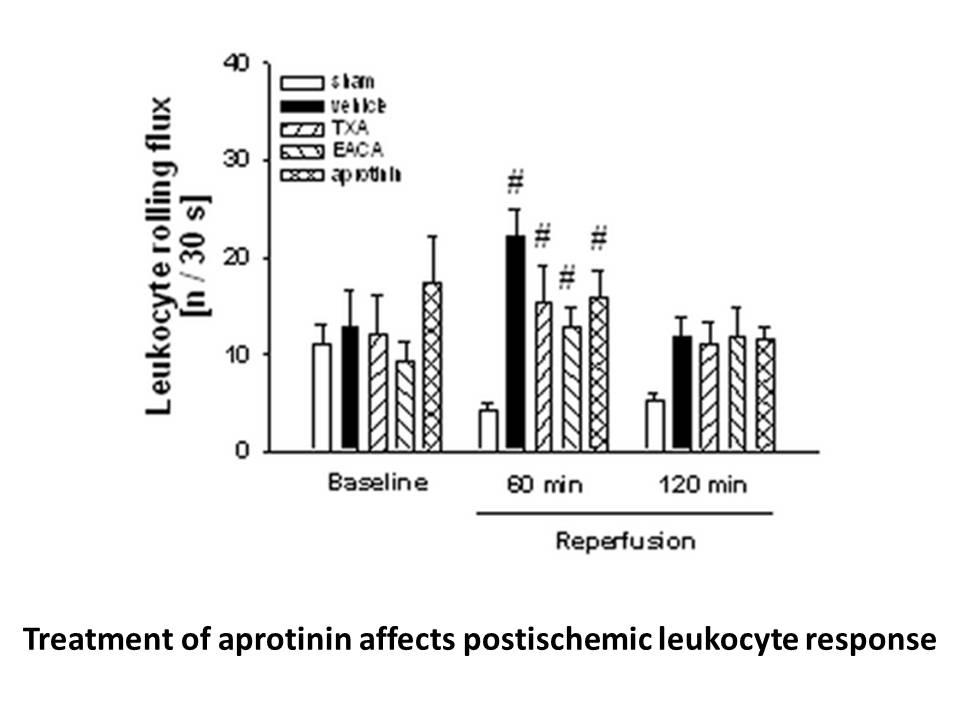
After ischemia, expression of transcription factors, including products of immediate early genes, stress proteins and neurotrophic factors are also altered in CA1 neurons78, 79. These proteins are potential candidates for downregulating of GluR2 expression by reducing mRNA transcription or stability
-
The bioconversion of inositol from
2022-03-03

The bioconversion of inositol from glucose was anticipated nearly a century ago and later confirmed by isotope tracing methods [[22], [23]]. This biosynthesis of inositol involves three sequentially acting enzymes: Firstly, ATP-dependent glucokinase converts glucose to glucose-6-phosphate (G-6P) [[2
-
It was previously proposed that glucagon acts
2022-03-02
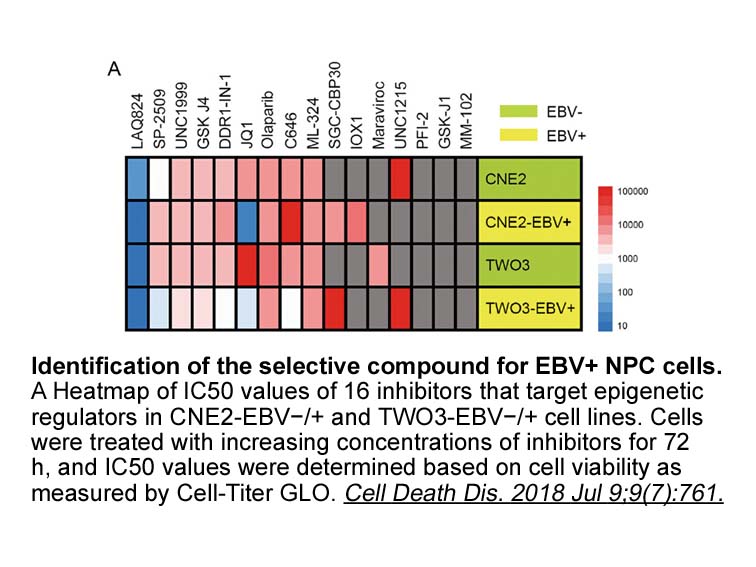
It was previously proposed that glucagon acts in the liver, in which a signal is produced and relayed to the Aurora Kinase Inhibitor III via vagal nerves [8], [36], [37]. The concept that the liver is the primary target site was supported by the studies reporting that the glucagon receptor is local
-
Extending time action is another strategy to increase
2022-03-02
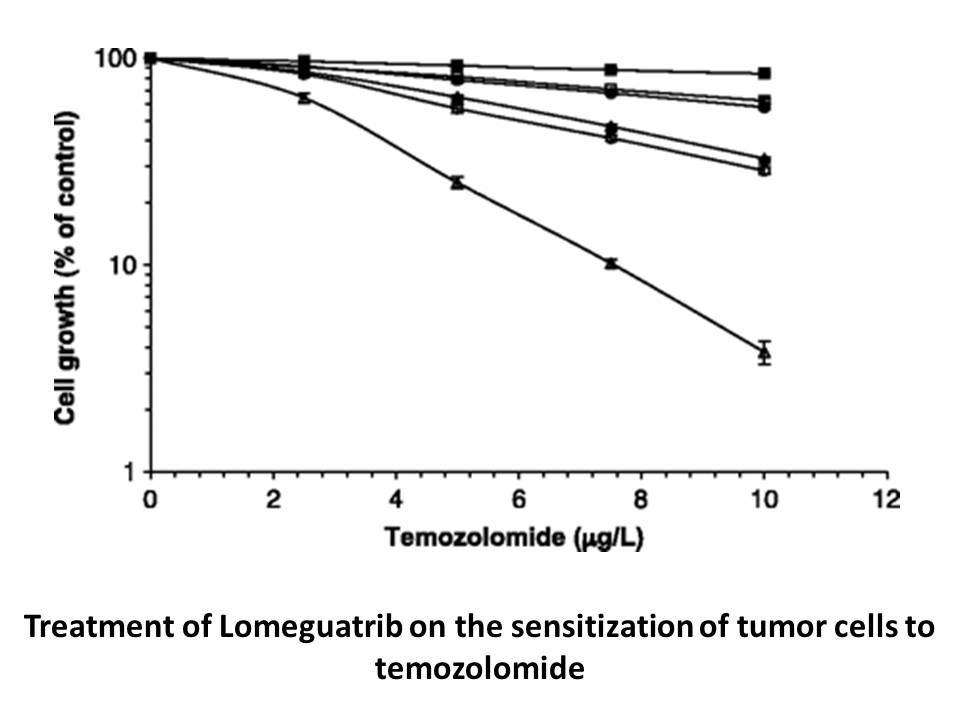
Extending time action is another strategy to increase in vivo efficacy. Twice daily administration of the unacylated analogs herein caused greater body weight loss in DIO mice than the equivalent dose administered as a single daily injection, suggesting that protracted time action should yield more
-
Species specificity of small molecular GPR agonists
2022-03-02
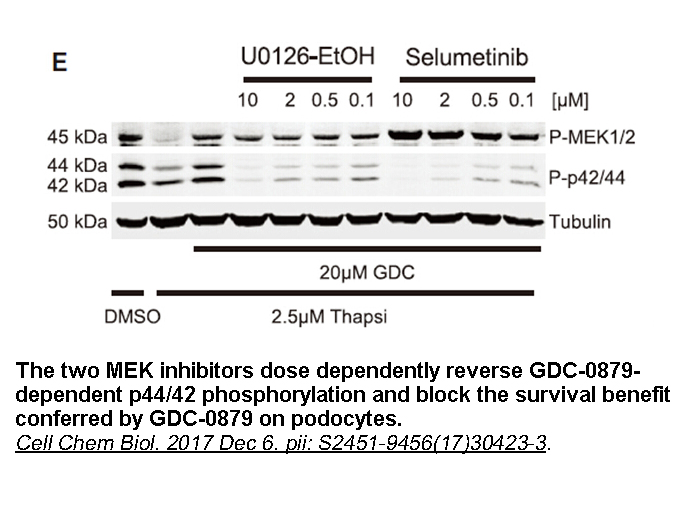
Species specificity of small molecular GPR40 agonists have been reported by Takeda scientist. In the binding pocket of TM5, a Leu186 in human GPR40/FFA1 is replaced with Phe in rat, resulting in dramatic inter-species GPR40 activity discrepancy in certain scaffolds of small molecular GPR40 agonists.
-
The pharmacokinetic properties of were amenable to
2022-03-02
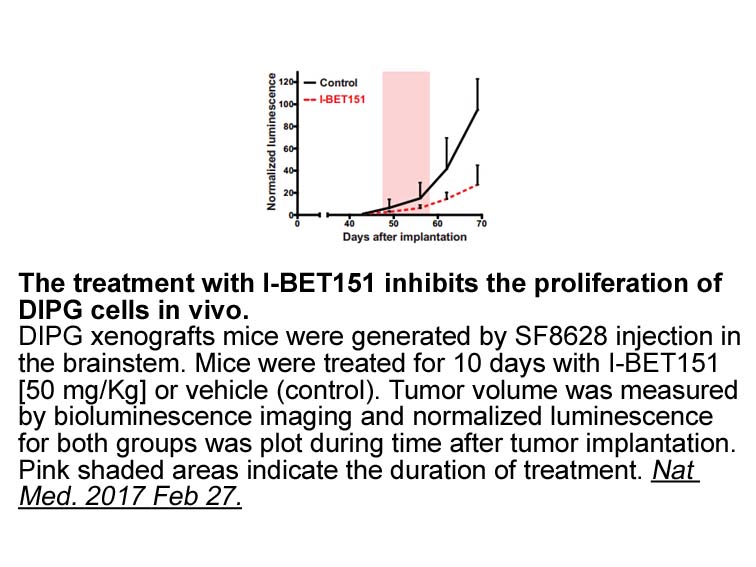
The pharmacokinetic properties of 12 were amenable to oral dosing allowing in vivo comparison to 6. Compared to a maximum efficacious dose of 6 (60mg/kg), 12 demonstrated improved glycemic control during OGTT in high-fat fed/STZ treated (HF/STZ) and BDF/diet-induced obesity (DIO) mouse models of typ
-
Magtanong et al observed that caspase
2022-03-02
Magtanong et al. (2019) observed that caspase-dependent apoptotic lipotoxicity of saturated fatty acids is suppressed by exogenous MUFAs. This may explain the ability of MUFAs to prevent atherosclerosis, in contrast to the pathogenicity of saturated fatty acids. Moreover, MUFAs may promote while PUF
-
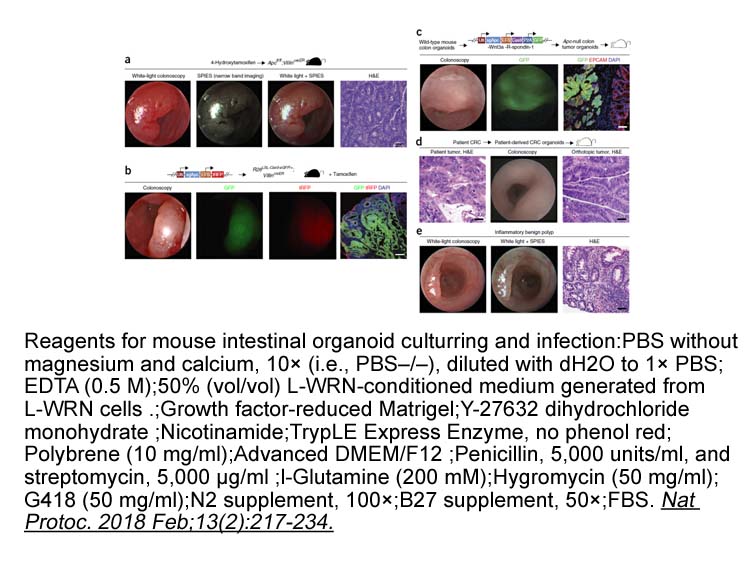
br Introduction The brain contains abundant fatty acids whic
2022-03-02
Introduction The brain contains abundant fatty acids, which serve as constituents of membranes and as an energy source. In addition, fatty acids and their metabolites contribute to signal transduction between neurons or neurons and glial cells, suggesting that fatty acids play a crucial role in d
-
Genetic contributions to this dysregulation
2022-03-02

Genetic contributions to this dysregulation of expression have been studied, and significant associations have been found. The Fas −670 polymorphism is associated with preterm premature rupture of membranes, preeclampsia, as well as intrauterine growth restriction [7], [16]. The Fas SNP (−670 A>G) i
-
ras inhibitor To validate that our natural experiment operat
2022-03-02
To validate that our natural experiment operates primarily through a large negative shock to the risk-taking incentives provided by option compensation measured by vega, we do two sets of tests. First, we confirm a significant drop in vega after the adoption of FAS 123R for treated firms relative to
-
Hyperactivation of Notch pathway can give rise
2022-03-02
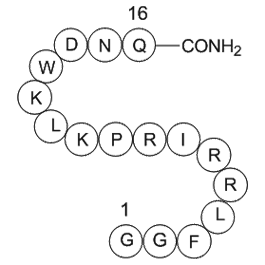
Hyperactivation of Notch1 pathway can give rise to transformation into T-ALL in murine models. In more than 50% of acute T-cell lymphoblastic leukemias oncogenic activating mutations of Notch1 have been detected and inactivation of Notch1 signaling impairs leukemia cell proliferation and promotes ap
-
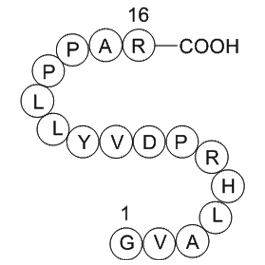
SB 204070 The mechanism study demonstrated that BDNF
2022-03-02
The mechanism study demonstrated that BDNF is one of the fundamental mechanisms underlying JMJD2A effects during neuropathic pain. BDNF is a neurotrophin with essential functions in neuronal development and neuroplasticity. Accumulating evidence suggests that alterations in BDNF expression levels un
-
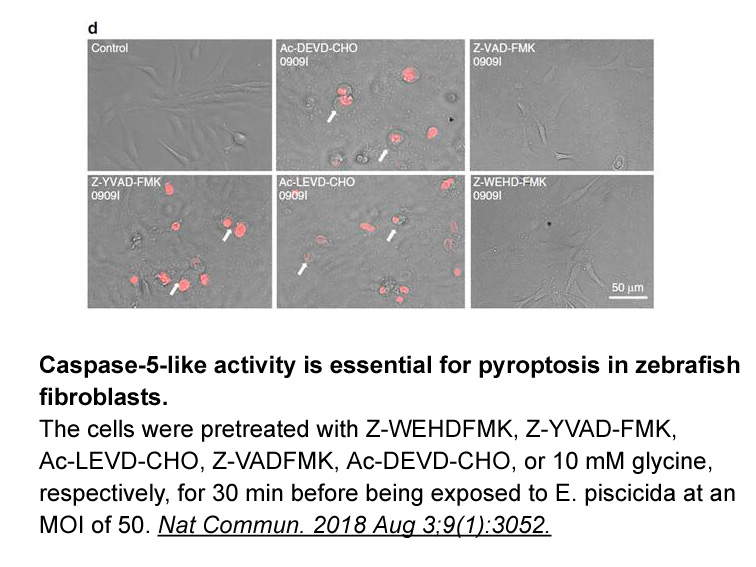
Intracellular signaling pathways leading to IR injury are
2022-03-02
Intracellular signaling pathways leading to IR injury are initiated by an increase in reactive oxygen species (ROS). This occurs principally at the time of initiation of reperfusion, which is accompanied by the re-admission of oxygen to the ischemic tissue. The resulting cascade of downstream events
-
G007-LK Hh signaling pathway plays an essential role in
2022-03-02
Hh signaling pathway plays an essential role in the regulation of invertebrate and vertebrate tissue development and metabolic homeostasis. Components of Hh signaling pathway have been identified in succession, including Hh ligands, membrane protein receptors, cytoplasmic components and transcriptio
-
Genetic disruption or pharmacologic inhibition
2022-02-28

Genetic disruption or pharmacologic inhibition of the hepatic glucagon pathway has invariably been shown to increase pancreatic α cell mass. This has been observed in glucagon receptor (GCGR) knockout (Gcgr−/−) mice (Gelling et al., 2003), glucagon knockout mice (Hayashi et al., 2009), prohormone co
15987 records 475/1066 page Previous Next First page 上5页 471472473474475 下5页 Last page